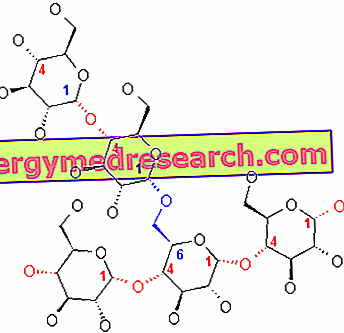
Le glycogène est une macromolécule (masse moléculaire d'environ 400 millions de daltons) de l'α-glucose dans laquelle se trouvent principalement des liaisons α-1, 4 glycosidiques et des ramifications dans un rapport de 1:10, en raison des liaisons α-1, 6 glycosidiques.
Le glycogène est un matériau de réserve qui est continuellement dégradé et reconstitué; Il y a environ 100 g de glycogène dans toute la masse cellulaire du corps: la plus grande partie se trouve dans le foie où il est mobile et peut donc être utilisé comme réserve pour les autres organes (le glycogène dans les muscles n'est pas mobile).
Les enzymes qui catalysent la dégradation et la synthèse du glycogène se trouvent toutes dans le cytoplasme. Nous avons donc besoin d’un système d’ajustement permettant de rendre l’un inactif inactif lorsque l’autre est actif: s’il existe une disponibilité de glucose, ce dernier est converti en glycogène. (anabolisme) qui est une réserve, et inversement, s’il existe une demande de glucose, le glycogène est dégradé (catabolisme).
L'enzyme principalement impliqué dans la dégradation du glycogène est la glycogène phosphorylase ; cette enzyme est capable de cliver une liaison α-1, 4 glycosidique en utilisant comme agent lytique un orthophosphate inorganique: la scission a lieu par voie phosphorolytique et le glucose 1-phosphate est obtenu.
À cinq ou six unités d'un point de ramification, l'enzyme glycogène phosphorylase n'est plus en mesure d'agir. Par conséquent, elle se détache du glycogène et est remplacée par une enzyme déramifiante qui est une transférase : dans le site catalytique de cette enzyme, histidine qui permet le transfert de trois unités saccharides vers la chaîne glycosidique la plus proche (l'histidine attaque le premier carbone d'une molécule de glucose). L'enzyme que je viens de mentionner est la glycosyltransférase ; à la fin de l'action de cette enzyme, il ne reste qu'une seule unité de glucose avec le premier carbone lié au sixième carbone d'un glucose de la chaîne principale. La dernière unité de glucose de la chaîne latérale est libérée par l'action de l'enzyme α-1, 6 glycosidase (cette enzyme constitue la deuxième partie de l'enzyme déramificante); puisque les ramifications dans le glycogène sont dans un rapport de 1:10, nous obtenons environ 90% de glucose 1-phosphate et environ 10% de glucose à partir de la dégradation complète de la macromolécule.
L'action des enzymes susmentionnées permet d'éliminer une chaîne latérale de la molécule de glycogène; l'activité de ces enzymes peut être répétée jusqu'à la dégradation complète de la chaîne.
Considérons un hépatocyte; Le glucose (assimilé par le régime alimentaire), lorsqu'il entre dans la cellule, est converti en glucose 6-phosphate et est donc activé. Le glucose 6-phosphate, sous l'action de la phosphoglucomutase, est transformé en glucose 1-phosphate: ce dernier est un précurseur non immédiat de la biosynthèse; dans la biosynthèse, on utilise une forme activée de sucres qui est représentée par un sucre lié à un diphosphate: généralement l’uridyldiphosphate (UDP). Le glucose 1-phosphate est ensuite converti en UDP-glucose; ce métabolite passe sous l'action de la glycogène synthase capable de lier l'UDP-glucose à une extrémité non réductrice du glycogène en croissance: on obtient le glycogène allongé d'une unité glucosidique et de l'UDP. L'UDP est convertie par l'enzyme nucléosidedifosfochinasi en UTP qui retourne à la circulation.
La dégradation du glycogène se produit sous l'action de la glycogène phosphorylase, qui libère une molécule de glucose et la transforme en glucose 1-phosphate. Ensuite, la phosphoglucomutase convertit le glucose 1-phosphate en glucose 6-phosphate.
Le glycogène est synthétisé avant tout dans le foie et les muscles: dans l'organisme, il y a 1 à 1, 2 hectolitres de glycogène répartis dans la masse musculaire.
Le glycogène d'un myocyte représente une réserve d'énergie uniquement pour cette cellule, tandis que le glycogène contenu dans le foie sert également de réserve pour d'autres tissus, c'est-à-dire qu'il peut être envoyé, comme le glucose, à d'autres cellules.
Le glucose 6-phosphate obtenu dans les muscles suite à la dégradation du glycogène est ensuite envoyé, en cas de besoin en énergie, à la glycolyse; dans le foie, le glucose 6-phosphate est converti en glucose par l'action de la glucose 6-phosphate phosphatase (enzyme caractéristique des hépatocytes) et est acheminé dans le courant circulatoire.
La glycogène synthase et la glycogène phosphorylase agissent toutes les deux sur les unités glycogènes non réductrices; il doit donc exister un signal hormonal qui commande l'activation d'un chemin et l'autre bloc (ou vice versa).
En laboratoire, il a été possible d’allonger la chaîne glycogène en exploitant la glycogène phosphorylase et en utilisant du glucose 1-phosphate à une concentration très élevée.
Dans les cellules, la glycogène phosphorylase ne catalyse que la réaction de dégradation car les concentrations des métabolites sont de nature à déplacer la balance de la réaction suivante vers la droite (c'est-à-dire vers la dégradation du glycogène):

Nous voyons le mécanisme d’action de la glycogène phosphorylase: l’oxygène acétal (qui fait office de pont entre les unités glucose) se lie à l’hydrogène phosphorylé: une réaction intermédiaire se forme donnée par un carbocatione (sur le glucose extrémité) à laquelle le phosphoryl (Pi) est lié très rapidement.
La glycogène phosphorylase nécessite un cofacteur qui est le phosphate de pyridoxal (cette molécule est également un cofacteur des transaminases): elle ne contient qu'un proton partiellement phosphorylé (le phosphate de pyridoxal est entouré d'un environnement hydrophobe qui justifie la présence de protons liés) . Phosphoryl (Pi) est capable de donner un proton au glycogène car ce phosphoryle réacquiert ensuite, le proton du phosphoryle partiellement protégé du phosphate de pyridoxal. La probabilité que, au pH physiologique, le phosphoryle perde le proton et reste complètement déprotoné est très faible.
Voyons maintenant comment fonctionne la phosphoglucomutase. Cette enzyme a un résidu sérine phosphorylé dans le site catalytique; la sérine donne phosphoryle en glucose 1-phosphate (en position six): pendant une courte période, le glucose se forme en 1, 6-bisphosphate, puis la sérine est réfrhosforilée en prenant le phosphoryl en position un. La phosphoglucus mutase peut agir dans les deux sens, c’est-à-dire convertir le glucose 1-phosphate en glucose 6-phosphate ou inversement; Si du glucose 6-phosphate est produit, il peut être directement envoyé à la glycolyse, dans les muscles, ou transformé en glucose dans le foie.
L'enzyme uridyl phosphoglucus transferase (ou UDP glucose pyrophosphorylase) catalyse la réaction de transfert glucose-1-phosphate en UTP en attaquant le phosphoryle a.
L'enzyme décrite ci-dessus est une pyrophosphorylase: ce nom est dû au fait que la réaction opposée à celle qui vient d'être décrite est la pyrophosphorylation.
Le glucose UDP, obtenu tel que décrit, est capable d’allonger la chaîne glycogène d’une unité monosaccharidique.
Il est possible de faire évoluer la réaction vers la formation de glucose UDP en éliminant un produit qui est le pyrophosphate; l'enzyme pyrophosphatase convertit le pyrophosphate en deux molécules d'orthophosphate (hydrolyse d'un anhydride) et maintient ainsi la concentration en pyrophosphate si basse que le processus de formation de glucose UDP est favorisé thermodynamiquement.
Comme mentionné précédemment, le glucose UDP, grâce à l'action de la glycogène synthase, est en mesure d'allonger la chaîne glycogène.
Les ramifications (dans un rapport de 1:10) sont dues au fait que, lorsqu'une chaîne glycogène est composée de 20-25 unités, une enzyme de ramification (ayant une istidine sur son site catalytique) intervient, capable de transférer une série de 7 -8 unités glycosidiques en aval de 5-6 unités: une nouvelle branche est ainsi générée.
Pour les questions d'origine nerveuse ou si de l'énergie est requise en raison d'efforts physiques, l'adrénaline est sécrétée par les glandes surrénales.
Les cellules cibles de l'adrénaline (et de la noradrénaline) sont celles du foie, des muscles et du tissu adipeux (dans ce dernier se produit la dégradation des triglycérides et la circulation des acides gras: dans la mitochondrie, le glucose est ainsi produit 6 -phosphate, à envoyer à la glycolyse, tandis que dans les adipocytes, le glucose 6-phosphate est transformé en glucose par l’enzyme glucose 6-phosphate phosphatase et exporté dans les tissus).
Voyons maintenant les modes d'action de l'adrénaline. L'adrénaline va se lier à un récepteur situé sur la membrane cellulaire (des myocytes et des hépatocytes), ce qui détermine la traduction du signal de l'extérieur vers l'intérieur de la cellule. La protéine kinase est activée et agit simultanément sur les systèmes qui régulent la synthèse et la dégradation du glycogène:
La glycogène synthase existe sous deux formes: une forme déphosphorylée (active) et une forme phosphorylée (inactive); la protéine kinase phosphoryle la glycogène synthase et bloque son action.
La glycogène phosphorylase peut exister sous deux formes: une forme active dans laquelle une sérine phosphorylée est présente et une forme inactive dans laquelle la sérine est déphosphorylée. La glycogène phosphorylase peut être activée par l'enzyme glycogène phosphorylase kinase . La glycogène phosphorylase kinase est active si elle est phosphorylée et inactive si elle est déphosphorylée; la protéine kinase a pour substrat la glycogène phosphorylase kinase, c'est-à-dire qu'elle est capable de phosphoryler (et donc d'activer) cette dernière, qui à son tour active la glycogène phosphorylase.
Une fois que le signal d'adrénaline est terminé, il doit également mettre fin à son effet dans la cellule: les enzymes phosphatases interviennent ensuite sur l'espèce protéique.